Protein Phosphorylation; Not just a binary switch.
Written by Joris Frenz; Proteomics and Cancer cell signaling PhD researcher at the German Cancer Research Center, Heidelberg
Phosphorylation-Mediated Protein Regulation
Proteins, the molecular workhorses orchestrating cellular functions, exhibit an astonishing degree of complexity in their regulation. Beyond the linear sequence of amino acids that defines a protein, the dynamic landscape of post-translational modifications adds layers of sophistication to their functionality. Among these modifications, phosphorylation emerges as a pivotal regulatory mechanism, acting as a molecular switch that can toggle proteins between active and inactive states.
While single phosphorylation events are often thought to serve as binary on-off switches, many proteins undergo multiple phosphorylation events, creating a complex network of regulatory phospho-proteins. The dynamic and context-dependent nature of these phosphorylation events shapes cellular responses, orchestrating a symphony of molecular events that dictate crucial processes such as transcription, cell cycle progression, and signal transduction. In this complex realm of post-translational modifications, the synergistic or antagonistic effects of multiple phosphorylation events contribute to the regulatory mechanisms governing cellular physiology and pathology.
This article will highlight certain proteins with multiple phosphorylation sites, which interplay with each other, leading to a more sophisticated regulatory mechanism than a direct activation or inactivation of the protein.
- p53 – The guard that needs guidance:
One of the most studied tumor suppressor proteins is the transcription factor p53, widely known as the guardian of the genome. Besides safeguarding genomic stability, it has crucial roles in cell cycle progression, DNA repair, and apoptosis. Dysregulation of p53 function is implicated in a wide array of human cancers, highlighting its central importance in maintaining cellular integrity and preventing aberrant cell growth.
Given p53’s activity is of such fundamental importance for the cell, it is tightly regulated by multiple phosphorylation sites. Phosphorylation of Ser15 promotes acetylation of p53 lysine residues in the DNA binding and carboxy-terminal domains, which in turn inhibits ubiquitylation of the protein by MDM2 and subsequent degradation by the proteasome. Thereby, phosphorylation of Ser15 stabilizes p53[1]. Further, Ser15 also enhances phosphorylation of Ser9, Ser20, Ser46, and Thr18.
Interestingly, Ser20 and Thr18 phosphorylation were also found to be crucial for the release of p53 from MDM2[2], indicating that the differential phosphorylation of each of these phospho-sites is suitable to tightly control p53 stability and degradation.
The activity of p53 is further regulated by the phosphorylation of other residues. For example, Ser46 phosphorylation acts as a pro-apoptosis factor,[3] while phosphorylation of Ser392 influences mitochondrial translocation of p53, acting as a further apoptosis inducer in a transcription-independent manner[4].
|
Phospho-P53 (Ser15) Polyclonal antibody (28961-1-AP) |
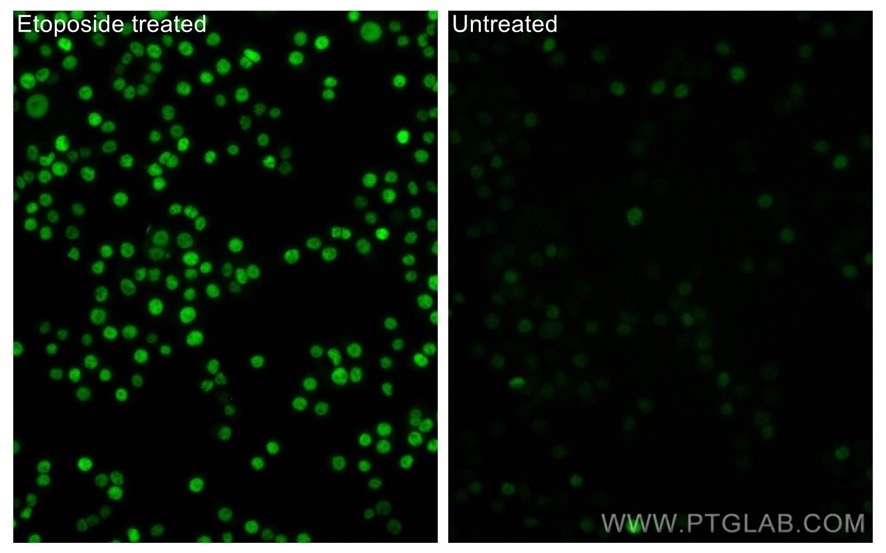 |
|
Immunofluorescent analysis of (4% PFA) fixed non-treated HT-29 and etoposide treated HT-29 cells using Phospho-P53 (Ser15) antibody (28961-1-AP) at dilution of 1:100 and CoraLite®488-Conjugated AffiniPure Goat Anti-Rabbit IgG(H+L). |
- Cyclin-Dependent Kinase 1 (CDK1) and the kinases cycling around it:
Cyclin-dependent kinase 1 (CDK1) is a pivotal regulator of the cell cycle, orchestrating the transition from the G2 phase to mitosis. Partnering with other cyclins, CDK1 forms complexes that drive key events in cell division, ensuring proper chromosomal segregation and cell proliferation. Its activity is tightly controlled to maintain precise cell cycle progression, making CDK1 a critical player in safeguarding genomic stability and governing cellular proliferation. Dysregulation of CDK1 is associated with various diseases, particularly cancer.
Thr161 phosphorylation by the cyclin H-CDK7 complex is essential for CDK1 activation, allowing it to drive cell cycle progression and other complex signaling networks. Simultaneously, inhibitory phosphorylation events at Tyr15 and Thr14 by Wee1 and Myt1 prevent premature entry into mitosis. Cdc25 phosphatase counteracts these inhibitory phosphorylation events, promoting CDK1 activation by dephosphorylating Tyr15 and Thr14.[5]
The balance between the activating (Thr161) and inhibitory (Tyr15/Thr14) phosphorylation sites determines CDK1’s functional state. The temporal coordination of kinase and phosphatase activity ensures that CDK1 activation is tightly linked to the cell cycle phase and prevents premature mitotic entry. This complex regulation goes beyond a simple on-off switch, enabling fine-tuned cellular responses to internal and external signals.
- Tau Protein – On the microtubules edge:
Tau is a microtubule-associated protein crucial for maintaining neuronal structure and function. Under normal conditions, tau stabilizes microtubules, supporting cellular transport. In neurodegenerative diseases, such as Alzheimer’s, abnormal tau phosphorylation disrupts its function, leading to the formation of neurofibrillary tangles and contributing to neuronal degeneration. Understanding tau’s role in microtubule dynamics is essential for unraveling the pathophysiology of neurodegenerative disorders and developing potential therapeutic interventions.
The regulation of tau binding to microtubules hinges on its phosphorylation status. Specifically, phosphorylation at Ser262 and Ser356, corresponding to neighboring microtubule-binding repeats, diminishes tau-microtubule interactions and reduces tau susceptibility to aggregation. Additionally, phosphorylation at residues outside the microtubule-binding domain (e.g., Ser214 and Thr231) decreases tau’s affinity for microtubules.
These findings imply that phosphorylation at distinct tau sites may exert contrasting effects on tau aggregation. Moreover, the interaction between the peptidyl-prolyl isomerase Pin1 and phosphorylated Thr231 facilitates PP2A binding in the trans configuration, inducing a conformational change that restores tau’s microtubule-binding ability.[6]
- RNA Polymerase II (RNA Pol II) – Stop and go in the mRNA traffic:
RNA Polymerase II (RNA Pol II) is a pivotal enzyme in gene expression, transcribing DNA into messenger RNA (mRNA). As the primary mediator of mRNA synthesis, RNA Pol II governs the intricate process of transcription initiation, elongation, and termination. Its accurate functioning is essential for the synthesis of diverse proteins, making RNA Pol II a central player in the regulation of cellular processes and the foundation of gene expression in eukaryotes.
Ser5 phosphorylation of RNA Pol II is crucial during initiation, promoting promoter clearance and recruitment of capping enzymes, crucial for mRNA stability. Once transcription is initiated, RNA Pol II pauses 25-50bp downstream of the transcriptional start site until the positive transcription elongation factor P-TEFb, composed of the cyclin-dependent kinase Cdk9 and the regulatory subunit Cyclin, is recruited. P-TEFb phosphorylates RNA Pol II, facilitating transcription elongation. The interplay of Ser5 and Ser2 is sequential and dynamic: Ser5 phosphorylation primes the polymerase for Ser2 phosphorylation. Moreover, phosphorylation can occur at Ser7 of the C-terminal domain, a modification associated with the transcription of small nuclear RNAs (snRNA), and the recruitment of the integrator complex.[7]
This orchestrated phosphorylation cascade ensures a coordinated and regulated transcriptional process, coupling initiation and elongation to finely control gene expression and mRNA production.
- AKT (Protein Kinase B) – An integrative AKTor:
AKT, also known as Protein Kinase B, is a key signaling protein central to cell survival, growth, and metabolism. Acting downstream of various receptors, AKT regulates diverse cellular processes such as protein synthesis, glucose metabolism, and apoptosis. Its intricate signaling network makes AKT a crucial mediator in physiological responses and a focal point in understanding aberrant cell signaling associated with conditions such as cancer and metabolic disorders.
Upon initiation of the signaling pathway, AKT undergoes phosphorylation at Thr308 by PDK1. This process involves AKT binding to phosphatidylinositol-3,4,5-triphosphate (PIP3), inducing a conformational change that exposes its kinase domain to PDK1 on the cell membrane. Maximal activation requires an additional phosphorylation at Ser473 by mTORC-2, significantly enhancing AKT activity. Together, phosphorylation at Thr308 and Ser473 serves as a marker for heightened AKT activity in response to growth factors. Subsequently, phosphorylated AKT orchestrates diverse biological actions by modifying downstream proteins, including members of the FoxO family, GSK-3β, and mTOR.[8]
The crosstalk between these two phosphorylation events integrates signals from different pathways, such as PI3K/AKT and mTOR, to finely regulate AKT activity and downstream cellular responses, including cell proliferation and survival.

Phosphorylation – Just the Prelude to Cellular Complexity:
The instances discussed emphasize the complexity of protein regulation through phosphorylation, showcasing the interplay of multiple events with synergistic or antagonistic effects on protein function. These examples span diverse signaling pathways, indicating the intricacy of known regulatory mechanisms. Yet, the complexity may extend even further, considering the vast array of cellular processes. Many additional examples, not covered here, contribute to the intricate landscape of phosphorylation-mediated regulation. Importantly, these regulatory mechanisms often involve an intricate interplay with various post-transcriptional modifications, regulatory proteins, cofactors, and other cellular molecules, introducing additional layers of complexity to cellular regulation and signaling.
Other examples of regulatory complex protein phosphorylations:
- Mitogen-activated protein kinases (MAPKs), such as ERK, JNK, and p38, are involved in cell signaling.
- MAPKs are activated through phosphorylation cascades, involving multiple phosphorylation sites, influencing their activity and cellular responses.
Glycogen Synthase Kinase 3 (GSK-3):
- GSK-3 is a serine/threonine kinase involved in various cellular processes.
- Phosphorylation at different sites, such as Ser9 for GSK-3β, regulates its activity, and this phosphorylation can be influenced by other upstream signaling pathways.
AMP-Activated Protein Kinase (AMPK):
- AMPK is a key sensor of cellular energy status.
- Phosphorylation at Thr172 is essential for AMPK activation, and multiple upstream kinases, including LKB1 and CaMKKβ, can influence this phosphorylation.
NF-κB (Nuclear Factor-kappa B) pathway:
- NF-κB is a transcription factor involved in immune and inflammatory responses.
- IκBα, an inhibitory protein bound to NF-κB, undergoes phosphorylation at Ser32 and Ser36. This phosphorylation marks IκBα for degradation, enabling NF-κB activation.
- The p65 subunit of NF-κB can undergo phosphorylation at Ser536 and Ser276, which are associated with enhanced transcriptional activity and nuclear localization.
Know your protein!
It is crucial to recognize that phosphorylation events are not always binary; thus, thorough research on your protein of interest is imperative before commencing your studies.
The following resources may prove beneficial for your studies:
Discover our range of phospho-antibodies against 140+ PTMs here:
View our interactive pathway posters below:
 |
 |
References
1 - Jayne Loughery, Miranda Cox, Linda M. Smith, David W. Meek, Critical role for p53-serine 15 phosphorylation in stimulating transactivation at p53-responsive promoters, Nucleic Acids Research, Volume 42, Issue 12, 8 July 2014, Pages 7666–7680, https://doi.org/10.1093/nar/gku501
2 - Nicolaou ST, Kannan S, Warwicker J, Verma CS. Activation of p53: How phosphorylated Ser15 triggers sequential phosphorylation of p53 at Thr18 by CK1δ. Proteins. 2022 Dec;90(12):2009-2022. doi: 10.1002/prot.26393. Epub 2022 Jul 14. PMID: 35752942; PMCID: PMC9796392.
3 - Liebl, M. C. and Hofmann, T. G. (2019). Cell fate regulation upon dna damage: p53 serine 46 kinases pave the cell death road. BioEssays, 41(12). https://doi.org/10.1002/bies.201900127
4 - Castrogiovanni, C., Waterschoot, B., De Backer, O. et al. Serine 392 phosphorylation modulates p53 mitochondrial translocation and transcription-independent apoptosis. Cell Death Differ 25, 190–203 (2018). https://doi.org/10.1038/cdd.2017.143
5 - Massacci, G., Perfetto, L. & Sacco, F. The Cyclin-dependent kinase 1: more than a cell cycle regulator. Br J Cancer 129, 1707–1716 (2023). https://doi.org/10.1038/s41416-023-02468-8
6 - Noble W, Hanger DP, Miller CC, Lovestone S. The importance of tau phosphorylation for neurodegenerative diseases. Front Neurol. 2013 Jul 1;4:83. doi: 10.3389/fneur.2013.00083. PMID: 23847585; PMCID: PMC3696910.
7 - Czudnochowski, N., Bösken, C. & Geyer, M. Serine-7 but not serine-5 phosphorylation primes RNA polymerase II CTD for P-TEFb recognition. Nat Commun 3, 842 (2012). https://doi.org/10.1038/ncomms1846
8 - Yingze Wei, Jianyun Zhou, Haiyan Yu, Xiaoxia Jin, AKT phosphorylation sites of Ser473 and Thr308 regulate AKT degradation, Bioscience, Biotechnology, and Biochemistry, Volume 83, Issue 3, 4 March 2019, Pages 429–435, https://doi.org/10.1080/09168451.2018.1549974

Related Content
Tips for detecting phosphoproteins by western blot
Post-translational Modifications: An Overview
Support
Newsletter Signup
Stay up-to-date with our latest news and events. New to Proteintech? Get 10% off your first order when you sign up.